Apuntes, resúmenes y trabajos para todos los estudiantes y cursos
Metabolismo del Carbono y Nitrógeno en Plantas
Fase Oscura de la Fotosíntesis
Es la fase formada por aquellas reacciones que no dependen de la luz, donde se produce la asimilación o fijación del carbono en forma de triosas, utilizando los productos de la fase clara. Esta fase puede ser llevada a cabo gracias al Ciclo de Calvin-Benson, el cual ocurre dentro del estroma.
Ciclo de Calvin-Benson
Es un ciclo independiente de la luz que utiliza ATP y NADPH obtenidos de la fase clara para asimilar CO2 proveniente del medio, que ingresa por los estomas, y se obtiene una triosa fosfato. Este ciclo se lleva a cabo en tres fases:
- Fijación del CO2
- Reducción
- Regeneración del Aceptor de CO2
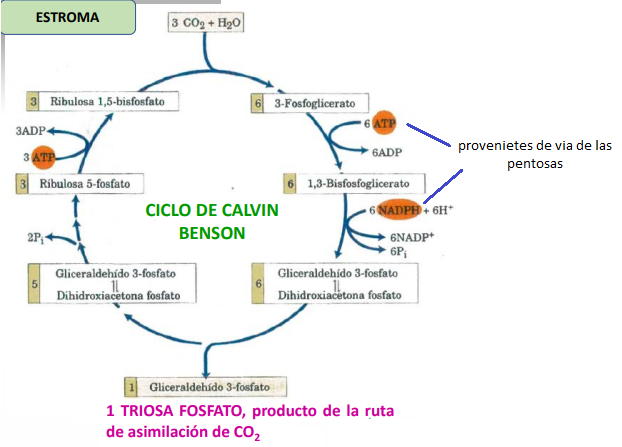
La primera reacción que se lleva a cabo es la Fijación de CO2, donde la ribulosa 1,5-bifosfato se carboxila añadiendo CO2 y luego se oxida para formar dos moléculas de 3-fosfoglicerato. Estas reacciones son catalizadas por la RUBISCO (Ribulosa-1,5-bifosfato carboxilasa/oxigenasa), dependiente de Mg+2.
Luego, este 3-fosfoglicerato sufre Reducción, donde se fosforila tras la lisis de un ATP y la acción de la fosfoglicerato quinasa, obteniendo 1,3-bifosfoglicerato, que por acción de la gliceraldehído 3-fosfato deshidrogenasa, la cual depende de la coenzima NADPH, se reduce a gliceraldehído 3-fosfato (TRIOSA FOSFATO).
Estas triosas tienen dos destinos: una de ellas sale del ciclo para la formación de almidón, y las otras cinco sufren una serie de reacciones de Regeneración de ribulosa 1,5-bifosfato, similares a las ocurridas en la vía de las pentosas, donde se producen interconversiones entre moléculas de azúcares fosfato que tienen de 3 a 7 carbonos, hasta llegar a ribulosa 5-fosfato. Esta, a través de la ribulosa 5-fosfato quinasa, regenera la ribulosa 1,5-bifosfato mediante fosforilación y la lisis de ATP, dando lugar nuevamente a la fijación de CO2.
El resultado de tres vueltas del ciclo es la conversión de 3 moléculas de CO2 y una molécula de fosfato en una TRIOSA FOSFATO. Por cada molécula de CO2 se produce un gliceraldehído 3-fosfato y se consumen 9 ATP y 6 NADPH obtenidos de la fase clara.
Todas estas reacciones del ciclo pueden ser llevadas a cabo en vegetales, pero en animales no todas. Las catalizadas por la RUBISCO, sedoheptulosa 1,7-bifosfatasa y la ribulosa 5-fosfato quinasa no se llevan a cabo en animales, por eso se dice que estos no pueden realizar la transformación neta de CO2 a glucosa.
RuBisCO en Carboxilación y Oxigenación
La RuBisCO tiene actividad carboxilasa y oxigenasa, es decir, puede tomar CO2 o también puede tomar O2 que provienen de la apertura de los estomas. Esta enzima posee regulación en cuanto a que es una metaloenzima dependiente del Mg+2.
Esta enzima también se regula por pH (ya que trabaja a pH óptimo), presencia de ATP, por disponibilidad de CO2. La luz también es un factor regulatorio de esta enzima. El 2-carboxiarabinitol-fosfato es un inhibidor, y también es regulada de forma alostérica por adición de un carboxilo al grupo amino de la subunidad grande, al cual se une Mg+2, activándola. La actividad oxigenasa es mayor a mayor temperatura. De esta manera, las plantas se han ido modificando para ser más eficientes ante condiciones adversas, y se diferencian en los siguientes grupos (se refiere a plantas C3, C4, CAM, aunque no se detallan aquí).
Destino de las Triosas Fosfato
Durante el Día
Durante el día, estas triosas obtenidas del ciclo de Calvin-Benson pueden ser usadas para:
- Gluconeogénesis Cloroplasmática: Formación de almidón.
- Gluconeogénesis Citoplasmática: Formación de sacarosa para metabolismo general.
Síntesis de Almidón
Es una vía cloroplasmática de las células fotosintéticas que se lleva a cabo durante el día para almacenar reservas para la noche. A partir de gliceraldehído 3-fosfato, el cual por acción de la gliceraldehído 3-fosfato isomerasa se transforma en dihidroxiacetona fosfato, ambos luego por acción de la aldolasa forman fructosa 1,6-bifosfato. Se realiza gluconeogénesis hasta formar glucosa 1-fosfato y, a partir de esta, por acción de la ADP-glucosa pirofosforilasa y la lisis de un ATP, se marca la glucosa y se forma ADP-glucosa, la cual se une a la estructura de almidón existente liberando ADP, por la almidón sintasa. Sobre esta estructura de almidón también actúa una enzima ramificante.
La regulación de este proceso se da a la altura de la ADP-glucosa pirofosforilasa, la cual se regula según la intensidad de la luz y la velocidad de la fotosíntesis. Durante la noche, el ATP se regenera muy lento y el Pi que se encuentra en el medio inhibe. El 3-fosfoglicerato formado en la fotosíntesis de día actúa como efector positivo.
Formación de Sacarosa (Citoplasma)
Parte de esa dihidroxiacetona fosfato sale fuera del cloroplasto mediante un cotransporte ubicado en la membrana interna del cloroplasto. Esta triosa sale fuera del cloroplasto y libera Pi. Cuando la concentración de Pi aumenta en el citoplasma, es signo de que la triosa fosfato fue utilizada. Entonces, este Pi vuelve a ingresar dentro del cloroplasto para formar ATP, hacer el ciclo de Calvin-Benson y formar nueva triosa, y así baja la concentración de Pi en el citoplasma y las triosas pueden salir fuera del cloroplasto. Por eso se lo llama sistema de transporte antiparalelo o cotransporte, porque sale triosa e ingresa Pi.
Luego de que la dihidroxiacetona fosfato se encuentra fuera del cloroplasto, puede ser utilizada para uso local (respiración) o para la formación de sacarosa. Dicho proceso parte de la unión de esta triosa y gliceraldehído 3-fosfato por acción de una aldolasa, dando como producto fructosa 1,6-bifosfato que por acción de la fructosa 1,6-bifosfatasa (enzima regulatoria) se transforma en fructosa 6-fosfato y realiza gluconeogénesis hasta formar glucosa 1-fosfato. Esta, por acción de la UDP-glucosa pirofosforilasa, dará UDP-glucosa. A través de la unión de esta última con la fructosa 6-fosfato por acción de la sacarosa-6-fosfato sintasa (enzima regulatoria alostérica), obtenemos sacarosa fosfato y luego por la sacarosa fosfato fosfatasa libera el Pi, dando como producto final SACAROSA, la cual posee un transportador y sale fuera de la célula a metabolismo general.
La regulación de este proceso se da a la altura de la fructosa 1,6-bifosfatasa y la sacarosa 6-fosfato sintasa, que posee regulación alostérica con glucosa 6-fosfato como efector positivo y Pi como efector negativo. Esta enzima se encuentra generalmente inactiva fosforilada; por una fosfatasa se desfosforila activándose ante una fotosíntesis activa (alta concentración de glucosa 6-fosfato) y se inactiva por una quinasa que la fosforila.
Durante la Noche
Durante la noche, al no ocurrir la fotosíntesis, se produce la degradación de ese almidón que fue formado durante el día para obtener dihidroxiacetona fosfato. Sobre este almidón actúan una serie de enzimas que son la almidón fosforilasa (libera glucosas por adición de fosfato dando glucosa 1-fosfato), la enzima desramificante (corta enlaces α 1-6 y forma dextrinas que por acción de las dextrinasas forman glucosa libre), las amilasas (cortan enlaces α 1-4 y forman maltosas que por acción de las α-glucosidasas forman glucosa libre). Esta glucosa libre es fosforilada en posición 6 por la hexoquinasa y la lisis de un ATP, transformándola en glucosa 6-fosfato. A partir de esta, realiza glucólisis hasta obtener gliceraldehído 3-fosfato y así transformarlo en dihidroxiacetona fosfato y sacarlo fuera del cloroplasto por el cotransporte y utilizarlo para formar sacarosa para metabolismo general y para uso propio por respiración.
Metabolismo del Nitrógeno
Transaminación
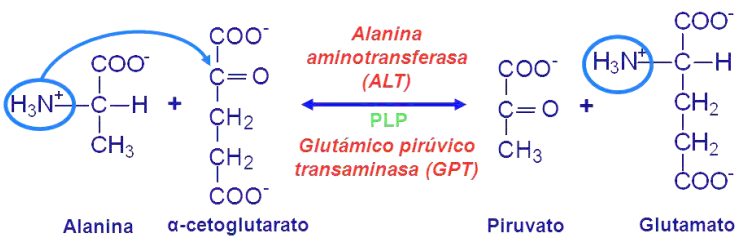
Es la transferencia reversible de un grupo amino de un aminoácido a un alfacetoácido, catalizada por una aminotransferasa usando piridoxal-fosfato (PLP) como coenzima, que proviene de las vitaminas de la dieta. El aminoácido se convierte en alfacetoácido y el alfacetoácido en el aminoácido correspondiente. El grupo amino no se elimina, sino que se transfiere.
Las principales reacciones de transaminación son:
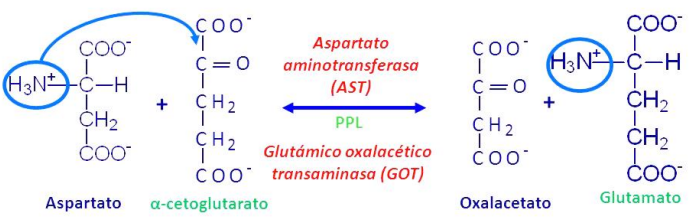
Esta reacción entre el aspartato y el alfacetoglutarato es muy importante en el hígado y corazón, en el citoplasma y mitocondria.
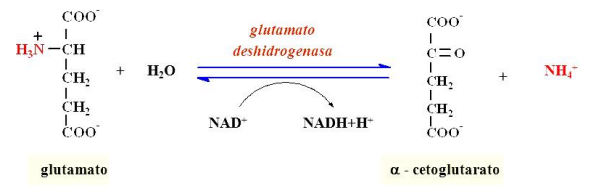
Desaminación
Como el alfacetoglutarato es uno de los principales sustratos de las transaminaciones, se produce la desaminación oxidativa del glutamato catalizada por la glutamato deshidrogenasa, la cual utiliza coenzimas NAD y NADP y forma alfacetoglutarato y amoníaco (NH3), el cual capta un protón y se transforma en NH4+ (amonio).
Eliminación de Amoníaco: Ciclo de la Urea
La eliminación de amoníaco es mediante el Ciclo de la Urea. Mediante este ciclo se produce la lisis de 4 enlaces de alta energía de 3 ATP y la eliminación de amoníaco en forma de urea. El ciclo se produce en la mitocondria (dos primeras reacciones) y en el citoplasma de los hepatocitos (tres reacciones). Esta urea se forma a partir de un ciclo que tiene 3 alimentadores: el amoníaco, CO2 y aspartato.
La primera reacción de este ciclo es la formación de carbamilfosfato mediante la condensación de CO2 (proveniente del ciclo de Krebs) y amoníaco (proveniente de la desaminación) mediante la lisis de 2 ATP por acción de la carbamilfosfato sintasa, dependiente de Mg+2 y N-acetilglutamato que actúa como activador. Luego se produce la síntesis de citrulina mediante la unión del carbamilfosfato formado anteriormente y ornitina mediante la ornitina transcarbamilasa. Luego se produce la adición del tercer alimentador, el aspartato, el cual se une a la citrulina y forma argininosuccinato mediante la argininosuccinato sintetasa, gracias a la lisis de dos enlaces de alta energía. Luego se libera fumarato, que tiene como destino el ciclo de Krebs, y la arginina continúa el ciclo para luego liberar urea por acción de la arginasa y formar ornitina para volver a hacer el ciclo. Los dos grupos amino de la urea provienen de la desaminación y del aspartato, y el carboxilo del CO2.
Fijación Biológica del Nitrógeno
Proceso de transformación del N2 atmosférico en amonio y su posterior fijación a una cadena carbonada gracias a un proceso biológico que ocurre dentro de las raíces de las LEGUMINOSAS por acción de una enzima que se denomina NITROGENASA y se encuentra presente en un MICROORGANISMO el cual se asocia con la planta y se lo llama RIZOBIO.
Para que la simbiosis entre la planta y el rizobio se lleve a cabo, deben transitarse una serie de etapas:
Etapas de la Simbiosis Rizobio-Leguminosa
- Intercambio de señales: La planta libera al suelo moléculas para interaccionar con los rizobios, llamadas flavonoides. Por su parte, la bacteria reconoce esa señal de la planta y responde con la liberación de una molécula llamada FACTOR NOD, los cuales son reconocidos por la planta y comienza el proceso de invasión de la bacteria dentro de la planta.
- Deformación del pelo radical: La bacteria se adhiere, la planta activa el ingreso, engloba las bacterias y forma una estructura conocida como cayado de pastor.
- Formación del hilo de infección: La bacteria comienza a ingresar por el pelo radical hasta la célula de la corteza externa o interna.
- Formación del nódulo: El hilo llega hasta la corteza externa o interna. Cuando llega a la corteza interna y posee células meristemáticas, se forma un nódulo alargado (plantas templadas, alfalfa). Cuando el hilo de infección llega a la corteza externa, se forman nódulos redondos sin células meristemáticas (plantas de climas más cálidos, soja, maní).
- Formación del simbiosoma dentro de la célula vegetal invadida: Una vez que la bacteria ingresa dentro de la célula a invadir, el rizobio se engloba con una membrana que forma el mismo hilo de infección al cerrarse. La bacteria comienza a ser un organismo que depende del carbono de la planta y se llama bacteroide. La membrana funciona como una barrera que controla el ingreso de oxígeno a esa célula.
Funcionamiento del Nódulo y Protección de la Nitrogenasa
La proteína leghemoglobina, producto de una simbiosis efectiva, capta O2 y sirve de barrera al mismo. Junto con la impermeabilidad de la membrana al O2 y la alta actividad metabólica del bacteroide, funcionan como barrera contra el O2 y permiten así el funcionamiento de la NITROGENASA.
Rizobacterias Promotoras del Crecimiento Vegetal (PGPR)
Las plantas liberan al suelo moléculas químicamente diversas que en conjunto se llaman exudados. Los microorganismos responden a ellos y se genera un tráfico molecular, creando una RIZOSFERA alrededor de la raíz. Estos microorganismos son de gran interés, ya que pueden ser benéficos, patógenos o de acción neutra para la planta, y al conjunto de los benéficos se los denomina PGPR (rizobacterias promotoras del crecimiento vegetal).
Mecanismos de Promoción de Crecimiento Directo
Existen mecanismos de promoción de crecimiento directo sobre la planta, como:
- Fijación de Nitrógeno: Transformación de N2 atmosférico en formas asimilables por la planta.
- Producción de Sideróforos: Moléculas que secuestran hierro, haciéndolo disponible para la bacteria y la planta, y limitando su disponibilidad para patógenos (en laboratorio se ve un halo anaranjado).
- Solubilidad de Fosfato: Bacterias capaces de solubilizar las sales de fosfato para que queden disponibles para las plantas (en el laboratorio se observa un halo transparente alrededor de la bacteria sembrada).
- Producción de Ácido Indolacético: Una auxina que estimula el crecimiento vegetal (en laboratorio se ve rojizo).